P2 — Map-guided foraging (the map's decay as survival pressure)
experiments/p2_map_guided_foraging.py
In Basal coupling the agent's HAP read whisker affordances only; food was found by the chance that wall-following crossed it. The map of the world existed, but the agent did not act on it — it was a picture for the modeller, not yet a tool for the agent.
This experiment closes that gap. The HAP now reads a second living-snapshot layer — a food memory — in which each consumption deposits a fading marker at the agent's dead-reckoned position. When the forward path is clear of walls and the food memory holds a live point in recall range, the HAP biases the turn toward the closest one. The map becomes for something the agent needs.
That turns the map's decay rate into a direct survival pressure: forget too fast → can't return to known food → starve under the same world.
Setup
Same body, balance beam, dead-reckoner, and living-snapshot wall map as Basal coupling. Added:
| component | role |
|---|---|
food_map |
a second OccupancyMap (its own decay rate, cap 3.0); consumption events deposit 2 hits at the dead-reckoned position, not the food's true coordinates |
| recall steering | when gate == 1 (path clear) and any live food-memory cell lies within [0.20, 2.0] m, turn := clip(2.0 · angular_error) toward the closest one |
Steering yields to wall avoidance whenever the HAP's standard (turn, gate)
asks for in-place rotation. The food memory does not override survival reflex.
The arena is harsher than basal coupling to make memory matter:
- 4 food items, mixed: 2 perimeter (
(0.8, 0.9),(-0.9, 0.7)— a wall-follower will find them) and 2 interior ((0.3, 0.2),(-0.4, -0.3)— only a remembering agent comes back). - Slow regrowth:
T_regrow = 90 s. - Long horizon:
T_end = 300 s(≈ 1.7× basal coupling). - Slightly richer food:
E_food = 32(the basal coupling used 25; the foraging task is harder, so each meal is worth more).
Conditions
| map use | food-memory decay | half-life | |
|---|---|---|---|
| C1 random | none | — | — (baseline) |
| C2 map-guided, slow | yes | 0.02 / s | ≈ 35 s |
| C3 map-guided, fast | yes | 1.00 / s | ≈ 0.7 s |
Everything else is identical (seed, body, biology, walls, food positions).
Result
C1 random (no map guidance) : died at t = 271.7 s, events = 5, mean E = 45, guided = 0.0%
C2 map-guided, slow decay (0.02/s) : alive at t = 300.0 s, events = 8, mean E = 60, guided = 21.3%
C3 map-guided, fast decay (1.0/s) : died at t = 175.1 s, events = 3, mean E = 28, guided = 0.3%
Three-way ordering, with the slow-memory agent the only survivor.
- C1 random dies at 272 s. The wall-following HAP visits the two perimeter foods reliably but never finds the interior ones — and two perimeter foods on a 90 s regrowth do not yield enough energy to cover the 300 s horizon.
- C2 map-guided, slow is alive at the end with 60% more consumption events than C1 (8 vs 5) and a mean energy 30% higher. The 21% of timesteps where the food memory steered the turn are concentrated near the interior locations the random forager never reaches.
- C3 map-guided, fast dies earlier than C1 (175 s vs 272 s). Fast decay is not merely useless — it is misleading: in the ~1 s window after a consumption the memory still pulls the agent back toward the just-eaten spot, where food will not regrow for another 90 s. The agent oscillates near recently-emptied spots, wastes energy, and dies sooner than the unguided baseline.
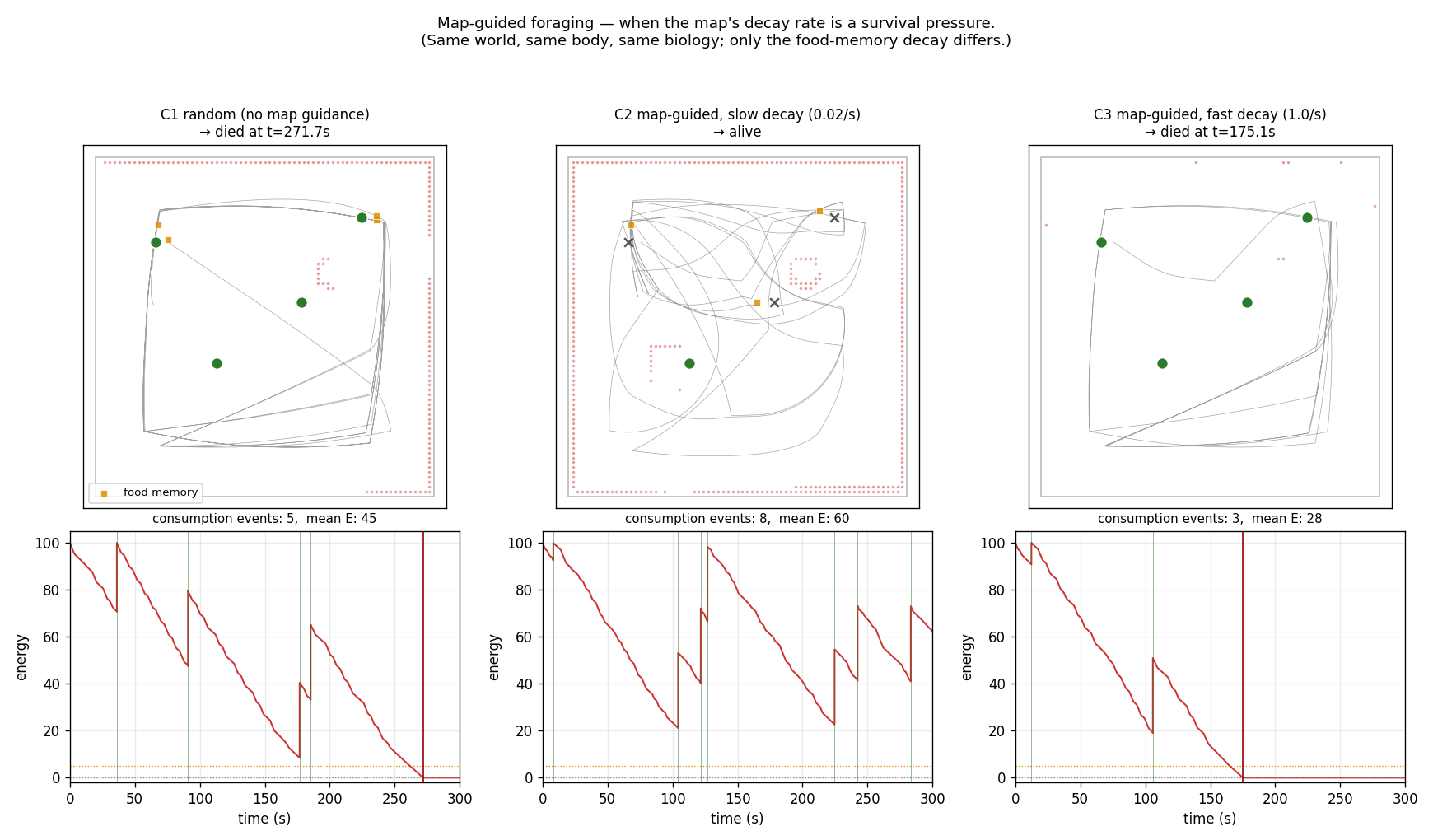
Top row per condition: trajectory + wall live map (red) + food memory live points (orange squares) + food (green = available, gray × = eaten). Bottom row: energy time-series with consumption (green) and death (red) markers. Only the slow-decay agent reaches the horizon alive; fast decay dies earlier than the no-map baseline.
What this experiment shows
It demonstrates the central claim from Basal coupling made quantitative:
- The same parameter that controls the cognitive substrate's perceptual fidelity (the living snapshot's decay rate) also controls survival. Slow decay → alive; fast decay → dead-sooner-than-no-map.
- Cognition does not sit on top of biology as a separable competence. The map is for finding food, food is for keeping the BAP alive, the BAP is for movement, and movement is for keeping the map. Sever any link — including by making the map fade too fast — and the loop opens.
- "Forgetting" is not neutral. Some decay is needed (otherwise the map hallucinates food at spots that have been emptied), but too much turns a reliable cognitive substrate into a misleading one. The optimum is set by the world's regrowth period — a property of the habitat, not of the agent.
It does not yet show:
- Quantitative robustness across seeds and parameter ranges (this is a single illustrative seed; the next refinement is a viability landscape sweep).
- Adaptive decay: the agent does not learn the world's regrowth period from experience. The decay rate is set, not discovered.
What it adds to the assumptions
Common assumptions hold, plus the Basal coupling biological state (energy reserve, energy-gated BAP, food as Python-tracked point items with regrowth). New:
- A food memory living-snapshot layer (its own
OccupancyMap), with its own decay rate, populated by consumption events (not by distance sensing). - The deposit is at the agent's dead-reckoned position, not the food's true coordinates — the memory lives in the agent's self-localized frame, so drift in dead-reckoning shifts the memory the same way it shifts the wall map. (See P2 self-localized for the drift budget.)
- A layered HAP: wall affordance > food memory > wander. Food-seeking is only invoked when the wall reflex is satisfied.
What is not added: distance sensing of food (the agent does not see food, it only feels it on contact); adaptive decay; multi-step planning. Each is a natural follow-up.
Why this experiment is in the bench
This is the first experiment in which the cognitive substrate is a survival organ, not just a representation. Basal coupling showed that movement sustains life through food; this one shows that remembering where food was is what makes that sustainability work — and that the quality of that memory (its decay rate) is a measurable survival pressure.
The same setup invites the obvious next sweeps:
- Decay × regrowth landscape — vary
(food_decay, T_regrow)and trace the viability boundary. The optimal decay should track the regrowth period (forget exactly when food becomes worth checking again). - HAP ablation under metabolism — does removing wall affordance, or the food-memory steering alone, collapse viability the way removing BAP did in the topology sweep?
- Adaptive decay — let the agent infer the regrowth period from inter-consumption intervals and adjust its own forgetting rate. A first primitive form of learning the habitat's dynamics.
Run it
cd experiments && ../.venv/bin/python p2_map_guided_foraging.py
Outputs: figures/p2_map_guided_foraging.png. Runtime: ~40 s on a laptop CPU.